

16S rRNA sequencing is a powerful molecular technique used to identify and classify bacteria and archaea based on their genetic material. It targets the 16S ribosomal RNA gene, which is present in all prokaryotes and contains both conserved and variable regions. These features make the 16S gene a reliable marker for distinguishing different microorganisms while allowing broad comparisons across taxa.
Several methodologies are commonly used for 16S rRNA sequencing. The traditional Sanger sequencing approach generates read lengths of approximately 700 to 1,000 base pairs, typically covering the full-length 16S rRNA gene. It provides high-quality, accurate results and is well established, making it useful for identifying single isolates or pure cultures. However, it is not practical for analyzing complex or mixed microbial communities such as those found in activated sludge systems. Consequently, this method is most often used in pure culture applications rather than community analysis.
Next-generation sequencing (NGS) platforms, such as Illumina (the most common) and Ion Torrent, are now widely used for microbiome and environmental studies. These methods produce shorter reads, typically 150–300 base pairs, although paired-end sequencing can extend this to around 600 base pairs. NGS offers high throughput and cost-effectiveness, but the shorter read lengths can limit taxonomic resolution. Commonly targeted regions of the 16S gene in Illumina-based sequencing include V1-V3, V3-V4 (the most popular), V4 and V4-V5.
More recently, third-generation (long-read) sequencing technologies such as PacBio SMRT and Oxford Nanopore have enabled sequencing of the full-length 16S gene (around 1,500 base pairs). These methods provide improved taxonomic resolution, often down to the species level, due to their long reads and full-gene coverage. However, they currently have higher error rates (which are steadily improving) and are more expensive than NGS approaches.
From a practical standpoint, it is important to recognize that not all DNA testing is the same, and different sequencing methodologies can produce different results. Understanding which method is used is essential when comparing data across studies or laboratories. Additionally, separate testing is often performed for bacteria and archaea. In anaerobic systems, for instance, methanogens are archaea, and both bacterial and archaeal analyses are recommended for a complete community assessment.
What’s actually measured in rRNA testing?
When interpreting rRNA testing results, it is important to understand that sequencing primarily measures relative DNA read percentages, which do not always directly correspond to the true abundance of organisms in a sample. A useful analogy is to think of microbes as having varying numbers of “fingers,” with rRNA representing those fingers. Counting DNA reads is like counting fingers — organisms with more “fingers” may appear more abundant even if their actual population is smaller.
Based on hundreds of DNA and microscopy split samples, there is often a weak correlation between DNA read percentage and the visible abundance of certain microorganisms. For example, as little as 2–3% of DNA reads from the genus Microthrix can correspond to abundant filamentous growth under the microscope. Conversely, genera such as Thauera, which often exhibit a zoogloea morphology, may register around 20% of total reads while showing only common to very common abundance when evaluated using the Jenkins 1-6 ranking scale. In some cases, general trends between DNA read percentages and microscopic abundance can be observed, but overall these two measures are not directly comparable in an “apples to apples” sense due to variability in rRNA copy number among different microbes.
Several other factors can also influence DNA read percentages. Microbes that grow rapidly tend to exhibit higher read percentages because fast-growing organisms often have multiple copies of rRNA genes. Similarly, organisms with high polysaccharide content generally show elevated read percentages. In contrast, slow-growing filamentous bacteria, such as those within the Chloroflexi phylum, may yield low read percentages despite appearing in high abundance under microscopic observation. For instance, it is common for only about 1% of Kouleothrix genus reads to correspond with very common to abundant Type 1851 filaments.
From a practical standpoint, DNA read percentages are best interpreted as indicators of “who’s there” rather than precise measures of abundance. Microscopy remains a valuable tool for assessing morphotype — that is, what the organisms look like and how abundant they appear. DNA sequencing complements microscopy by providing greater taxonomic detail and confirming the identity of observed organisms. Importantly, morphotype-to-cause correlations have held up strongly over more than 40 years of operational practice when samples are interpreted in the context of the surrounding microbial community.
DNA: Functional groups and morphology
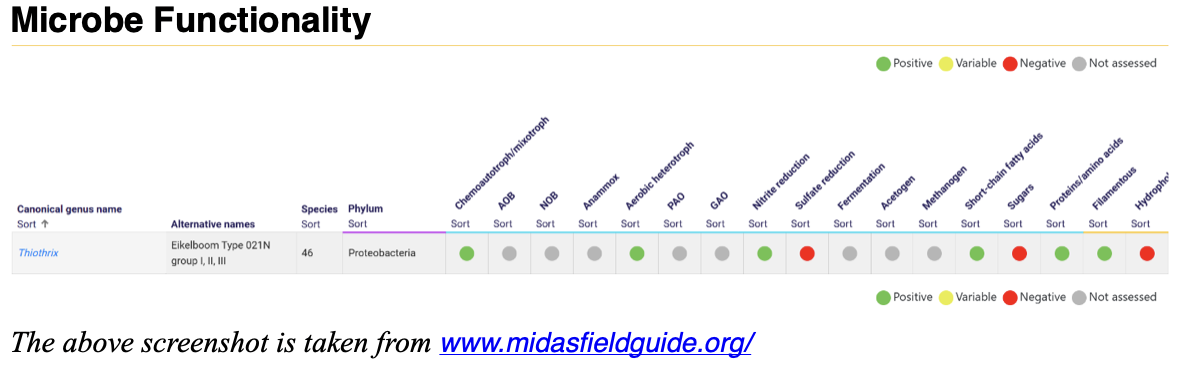
When referencing a genus in the MiDAS Field Guide, the entries typically include highlighted indicators for positive, variable, negative and unknown functionalities. It is important to understand that many genera have variable potential functionalities, meaning that just because a microorganism is capable of performing a certain function — often demonstrated under bench-scale conditions — this does not necessarily confirm that it is performing that role in situ within a biological wastewater treatment system.
Most genera capable of filamentous forms are labeled as “variable” because they may or may not exhibit filamentous morphology under the microscope, depending on environmental and operational conditions. Similarly, many genera display overlapping functional potentials, such as Candidatus Accumulibacter, which may act as both a polyphosphate-accumulating organism (PAO) and a glycogen-accumulating organism (GAO). These functional complexities are further amplified by the fact that numerous genera are capable of both oxidative and fermentative metabolic pathways. Even in cases such as enhanced biological phosphorus removal (EBPR), many of the involved genera may also possess the potential for denitrification.
DNA sequencing remains an emerging and rapidly advancing field, and the amount of available information varies widely between genera as of 2025. In practice, it is common for a portion of sequencing reads to remain unclassified at lower taxonomic levels, which is an issue that will be discussed in more detail later in this article.
What’s in a name?
It’s also good to recognize that there is no universally accepted database for microbial taxonomic classification. In practice, the MiDAS Field Guide is often considered the most practical and wastewater-relevant resource. However, several other credible and widely recognized databases are also in use, including:
• NCBI
• LPSN
• GTDB
• SILVA
• RDP
• EzBioCloud
• BacDive
Taxonomic assignments, genus names, and classifications may vary depending on which database is referenced, making it important to consider the context and source of the taxonomy when interpreting results.
On the microscopy side, there is generally broad genetic diversity within a single morphological description. For example, as illustrated in the referenced screenshot, there are 46 recognized species within the Thiothrix genus. This means that even within a single genus, not all organisms are genetically identical. Moreover, there are many closely related microbes capable of displaying a Thiothrix-like morphology while not actually belonging to the Thiothrix genus.

Basic information on taxonomy
The surrounding image is an example portion of a DNA report courtesy of Aster Bio. In this example the Oxford Nanopore platform is used with a database predominantly based around the MiDAS database, along with added information from other sources that have been deemed credible. Reads in these instances cover kingdom, phylum, class, order, family and genus, and read as low on the chart as accurately determined by the program. Blank spaces indicate that the margin of error was not within the designated range for confident classification at a particular taxonomic level.
DNA is an evolving field and as more knowledge is obtained occasional changes to classifications will occur. As technology is improving the criteria for genus classification in many instances is narrowing. It is important to not only consider the genus reads, but also higher taxonomic orders for instances in which functionality at these higher orders may capture a wider range of the big-picture conditions.
Many DNA reports on functional groups may be based on genus level reads, and it is important to take into consideration that there may be varying amounts of reads that do not reach the genus level.
There are some phyla in which general functionalities/morphotypes appear to be consistent (i.e., Chloroflexi as slower growing filamentous bacteria, many capable of storing substrate under anaerobic conditions) and also other phyla and taxonomic levels which may have significantly more diversity amongst functionality/morphology (i.e., Nitrosomonas and Thiothrix belong to the same phylum.)
Benefits and limitations of current DNA testing
Among the benefits of current DNA testing is that it is repeatable and non-subjective provided that known information about testing methods and databases are used. Also, databases are growing at a high rate in which more information is being learned about microbes at bench scale, in pure culture, and in situ.
While read percentage is not always a strong and reliable comparison to microbe abundance as viewed under the microscope, trends are effective in many instances depending on given objectives. Technology is advancing and pricing as well as turn-around time for DNA testing is becoming more practical and logistical.
Among its limitations are that DNA testing does not provide reliable context to conditions such as floc structure characteristics (i.e., firm vs. diffuse floc structure), dispersed growth abundance, and bacterial health/viability. Also, higher life form organisms and other organisms require different methodologies than community analysis for bacteria.
There is variable functionality and morphology amongst a high percentage of genus. DNA is an excellent tool for establishing if a microbe is present. However, if morphology (i.e., filamentous vs. non-filamentous) and functionality are variable, other tools such as microscopy and in-house analytical testing may be needed to help take this information into context. For example, Acinetobacter genus is commonly correlated with type 1863 filaments, however, doesn’t always display type 1863 filament morphology under the microscope).
Tools such as microscopy, in-house analytical testing, DNA testing and the intuition of operators familiar with treatment plant behavior are all relevant and useful tools. Gathering information from multiple sources of these tools gives us the higher levels of baseline data to use for troubleshooting and operational process control insight.
Tools and methodologies are only as good as the quality in which they are obtained. It is also important to know the context and the other potential variables for the best chances of practical application for these tools.
About the author: Ryan Hennessy is the principal scientist at Ryan Hennessy Wastewater Microbiology. He was trained and mentored by Dr. Michael Richard for over 10 years in wastewater microbiology, and serves as a microbiology services consultant. Hennessy is a licensed wastewater treatment and municipal waterworks operator in the state of Wisconsin and fills in as needed for operations at several facilities. He can be reached at ryan@rhwastewatermicrobiology.com. Hennessy's new book Wastewater Microbiology: Filamentous Bacteria Morphotype Identification Techniques, and Process Control Troubleshooting Strategies is now available on Amazon.












